


石斛属(Dendrobium S.W.)有1 200~1 500种,是兰科(Orchidaceae)中的大属之一,中国石斛属约有80种[1]。金钗石斛(Dendrobium nobile Lindl.)是该属中分布较广的物种,主要生长在热带、亚热带原始森林及类似的温暖湿润环境中,惧怕霜冻或高温[2-3]。金钗石斛是一种重要的药用草本植物,据中国古代医学书籍《本草纲目》记载,具有“强壮身体”的功效。金钗石斛以茎入药,性寒、味苦、淡、微咸,具有增强免疫力、强阴益精、生津养胃、润肺止咳、抗肿瘤等功效[4-5]。除药用外,金钗石斛花朵艳丽、花期较长,也具有较高的观赏和园艺价值[6]。近年来,大量采集和生境破坏导致金钗石斛野生资源稀缺,已被国家列入濒危植物名录而加以保护。如何合理开发利用和保护该重要的植物资源,是众多学者研究的方向,然而,作为研究基础的种质资源收集与评价,目前还没有系统的研究和报道。
1 材料与方法
1.1 实验材料
金钗石斛野生植株,共17个居群215个个体,其中我国云南省4个居群、广西壮族自治区4个居群、贵州省2个居群、四川省2个居群、西藏自治区2个居群、海南省1个居群,老挝1个居群、缅甸1个居群,采集后种植于南京师范大学植物学阳光房中。金钗石斛植物材料信息详见表1。
表1 金钗石斛17个居群的采集地点及样本数
Table 1
| 编码 | 采样点 | 样本数/株 | 经度E | 纬度N | 海拔/m |
|---|---|---|---|---|---|
| GXBS | 百色,中国广西壮族自治区 | 15 | 106°61' | 23°91' | 202 |
| GXJX | 金秀,中国广西壮族自治区 | 12 | 110°19' | 24°13' | 906 |
| GXBY | 宾阳,中国广西壮族自治区 | 13 | 108°80' | 23°22' | 117 |
| GXRX | 容县,中国广西壮族自治区 | 15 | 110°55' | 22°89' | 121 |
| YNBS | 保山,中国云南省 | 11 | 99°14' | 25°11' | 1 872 |
| YNHH | 红河,中国云南省 | 12 | 103°36' | 23°32' | 1 405 |
| YNLC | 临沧,中国云南省 | 15 | 100°06' | 23°87' | 1 760 |
| YNPE | 普洱,中国云南省 | 13 | 100°96' | 22°83' | 1 378 |
| GZCS | 赤水,中国贵州省 | 14 | 105°68' | 28°59' | 289 |
| GZLB | 荔波,中国贵州省 | 13 | 107°87' | 25°41' | 475 |
| XZCY | 察隅,中国西藏自治区 | 13 | 97°46' | 28°65' | 2 691 |
| XZMT | 墨脱,中国西藏自治区 | 15 | 95°33' | 29°33' | 1 008 |
| SCLS | 凉山,中国四川省 | 13 | 102°32' | 27°91' | 1 868 |
| SCXC | 西昌,中国四川省 | 16 | 101°73' | 27°20' | 2 162 |
| HNBS | 白沙,中国海南省 | 12 | 109°43' | 19°17' | 391 |
| LW | 老挝国 | 13 | 102°49' | 19°85' | 685 |
| MD | 缅甸国 | 15 | 95°84' | 22°24' | 617 |
1.2 实验方法
1.2.1 植株形态性状检测
于开花期检测金钗石斛植株性状。每个性状指标测定10个单株,求平均值,数值由 “平均值±标准差” 表示。用精度0.1 cm的直尺测量株高(cm)、叶片长(cm)、叶片宽(cm)、节间长(cm), 用精度0.1 mm的游标卡尺测量茎粗(mm), 叶片选取从顶叶向下第4片叶子,节间和茎粗测量部位选取从顶端向下第4节。
1.2.2 花性状检测
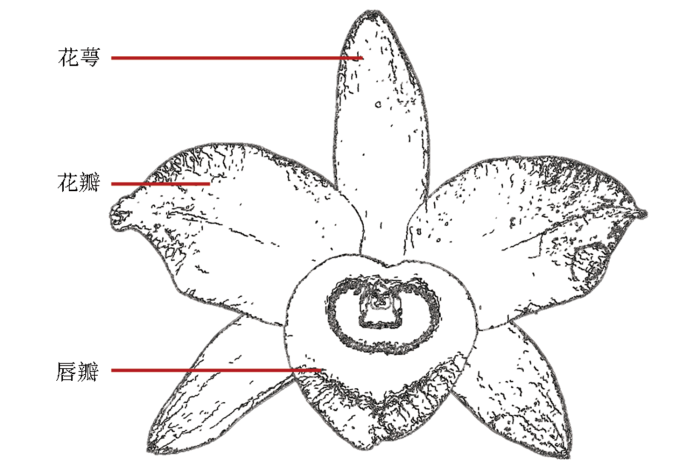
于盛花期检测金钗石斛花性状指标。同样,每个指标测定10个单株,求平均值,数值由“平均值±标准差”表示。用精度0.1 cm的直尺测量花径(cm)、花梗长(cm)、唇瓣长(cm)、唇瓣宽(cm)、花瓣长(cm)、花瓣宽(cm)、萼片长(cm)、萼片宽(cm),花结构如图1所示。
图1
1.2.3 气孔的显微结构观察
选取金钗石斛上部成熟叶片,用蒸馏水洗净叶片下表面,用镊子撕取叶片下表面中部主脉附近的表皮(0.5 cm×0.5 cm)放到载玻片上,再在表皮上加1滴蒸馏水,盖上盖玻片,用吸水纸在盖玻片上轻压吸取多余的水分。在10×10倍显微镜下,计5个视野的气孔数量,在10×40倍显微镜下,测5个保卫细胞的长和宽,取均值。计算:气孔比=气孔长度/气孔宽度,气孔面积=气孔长度×气孔宽度×3.14×0.25,气孔密度=气孔数/视野面积。
1.2.4 有效成分检测
(1)总生物碱提取与检测。将金钗石斛茎切成薄片,在60℃的烘箱中干燥36~48 h至绝对干燥后,测量干重(DW,g/L);精确称取0.5 g烘干后材料,在研钵中使用石英砂研磨成粉末,添加氨水溶液,混合物静置0.5 h。将混合液体倒入50 mL烧瓶中,并添加25 mL氯仿,在70℃水浴中保持2.5 h后进行萃取,再用旋转蒸发器干燥,然后向烧瓶中倒入5 mL氯仿以溶解干残渣,吸取2 mL氯仿提取物,并将氯仿添加到提取物中至10 mL。将邻苯二甲酸氢钾缓冲液(5 mL,pH 4.5)和2 mL 0.04%(m/V)溴甲酚绿溶液混合并倒入氯仿提取物中,剧烈摇动混合物3 min,静置30 min,向5 mL低馏分中加入1mL碱性乙醇(0.01 mol/L NaOH)进行分析。使用分光光度计在620 nm处测定吸光度值,然后使用制定的标准曲线方程计算总生物碱含量。使用以下公式计算总生物碱含量:C总生物碱(mg/g DW)=石斛碱(mg)×5/0.5 g [10]。
(2)多糖的提取与检测。将金钗石斛茎低温烘干并磨成粉末,准确称取0.10 g粉末置于100 mL三角瓶中,加沸蒸馏水25 mL,加盖,沸水浴中提取30 min,冷却后用滤纸过滤,残渣用沸蒸馏水反复洗涤后过滤,滤液收集在50 mL容量瓶中,用蒸馏水定容后即为提取液;采用硫酸-蒽酮法测定多糖含量[11],首先吸取提取液0.5 mL放入干净的试管中,加入1.5 mL蒸馏水,摇匀,然后依次加入0.5 mL蒽酮试剂和5 mL浓硫酸,显色后在620 nm波长下测定吸光度值,最后使用制定的标准曲线方程计算多糖含量。
1.2.5 数据处理 数据统计分析由SPSS 22.0软件完成。
2 结果与分析
2.1 植株形态性状分析
17个居群的金钗石斛植株形态如图2所示、表观指标数据如表2所示。从不同居群的金钗石斛形态特征可以看出,老挝(LW)居群叶片最长[(11.69±1.04) cm],贵州荔波(GZLB)居群叶片最短[(7.48±0.62) cm];云南宝山(YNBS)居群叶片最宽[(3.02±0.37) cm],西藏墨脱(XZMT)居群叶片最窄[(1.54±0.11) cm];植株最高的是缅甸(MD)居群[(44.17±3.86) cm],最矮是广西金秀(GXJX)居群[(15.25±1.36) cm];节间最长也是缅甸(MD)居群[(4.09±0.37) cm],最短是西藏墨脱(XZMT)居群[(2.03±0.15) cm];云南宝山(YNBS)居群茎粗最大[直径为(12.82±1.18) mm],西藏墨脱(XZMT)居群茎粗最小[直径为(6.30±0.52) mm]。
图2

表2 金钗石斛17个居群的形态数据
Table 2
| 编码 | 叶片长/cm | 叶片宽/cm | 株高/cm | 节间长/cm | 茎粗/mm |
|---|---|---|---|---|---|
| GXBS | 10.32±1.07 | 2.43±0.18 | 20.89±1.49 | 3.31±0.22 | 9.33±0.91 |
| GXJX | 8.38±0.60 | 2.41±0.21 | 15.25±1.36 | 2.58±0.18 | 9.86±0.61 |
| GXBY | 11.42±1.06 | 1.96±0.18 | 29.47±2.17 | 3.67±0.10 | 6.89±0.52 |
| GXRX | 7.63±0.34 | 2.23±0.31 | 15.86±1.44 | 2.88±021 | 7.74±0.33 |
| YNBS | 11.54±1.06 | 3.02±0.37 | 31.38±2.94 | 3.93±0.34 | 12.82±1.18 |
| YNHH | 11.09±0.67 | 2.29±0.19 | 28.26±2.62 | 2.90±0.19 | 10.94±1.07 |
| YNLC | 10.85±0.49 | 2.57±0.23 | 30.62±2.74 | 3.38±0.28 | 10.70±1.50 |
| YNPE | 8.91±0.73 | 1.98±0.13 | 19.26±1.25 | 2.38±0.19 | 8.60±0.87 |
| GZCS | 7.77±0.51 | 1.82±0.12 | 21.38±1.29 | 3.30±0.32 | 8.03±0.59 |
| GZLB | 7.48±0.62 | 1.71±0.13 | 38.63±2.63 | 3.89±0.12 | 11.05±1.11 |
| XZCY | 8.79±0.50 | 2.27±0.10 | 17.13±1.42 | 3.49±0.22 | 8.88±0.42 |
| XZMT | 7.62±0.63 | 1.54±0.11 | 30.55±2.28 | 2.03±0.15 | 6.30±0.52 |
| SCLS | 10.02±1.16 | 2.21±0.09 | 16.02±1.41 | 3.14±0.12 | 9.03±0.96 |
| SCXC | 7.95±0.51 | 1.84±0.15 | 19.68±1.55 | 2.89±0.32 | 11.76±1.15 |
| HNBS | 10.16±0.93 | 2.04±0.18 | 40.63±4.46 | 3.09±0.20 | 8.08±0.51 |
| LW | 11.69±1.04 | 2.72±0.18 | 39.39±3.50 | 3.32±0.16 | 12.71±0.85 |
| MD | 9.37±0.88 | 1.85±0.12 | 44.17±3.86 | 4.09±0.37 | 10.43±0.92 |
变异系数是反映数据离散型的指标,变异系数越小、数值的离散度越小。17个居群的金钗石斛植株表观指标数据的变异系数如表3所示,不同表观指标数据的平均变异系数不同,其中节间长的平均变异系数最小,为6.87,叶片长和叶片宽次之,分别为7.85和7.95,第三为株高,为8.26,茎粗的变异系数最大,为8.44。说明节间长和叶片长宽的性状稳定性比茎粗和植株高度更好,可作为优选性状指标应用于金钗石斛表观性状的评价中。
表3 金钗石斛17个居群的形态数据的变异系数
Table 3
| 编码 | 叶片长 | 叶片宽 | 株高 | 节间长 | 茎粗 |
|---|---|---|---|---|---|
| GXBS | 10.37 | 7.41 | 7.13 | 6.65 | 9.75 |
| GXJX | 7.16 | 8.71 | 8.92 | 6.98 | 6.19 |
| GXBY | 9.28 | 9.18 | 7.36 | 2.72 | 7.55 |
| GXRX | 4.46 | 13.90 | 9.08 | 7.29 | 4.26 |
| YNBS | 9.19 | 12.25 | 9.37 | 8.65 | 9.20 |
| YNHH | 6.04 | 8.30 | 9.27 | 6.55 | 9.78 |
| YNLC | 4.52 | 8.95 | 8.95 | 8.28 | 14.02 |
| YNPE | 8.19 | 6.57 | 6.49 | 7.98 | 10.12 |
| GZCS | 6.56 | 6.59 | 6.03 | 9.70 | 7.35 |
| GZLB | 8.29 | 7.60 | 6.81 | 3.08 | 10.05 |
| XZCY | 5.69 | 4.41 | 8.29 | 6.30 | 4.73 |
| XZMT | 8.27 | 7.14 | 7.46 | 7.39 | 8.25 |
| SCLS | 11.58 | 4.07 | 8.80 | 3.82 | 10.63 |
| SCXC | 6.42 | 8.15 | 7.88 | 11.07 | 9.78 |
| HNBS | 9.15 | 8.82 | 10.98 | 6.47 | 6.31 |
| LW | 8.90 | 6.62 | 8.89 | 4.82 | 6.69 |
| MD | 9.39 | 6.49 | 8.74 | 9.05 | 8.82 |
| 均值 | 7.85 | 7.95 | 8.26 | 6.87 | 8.44 |
2.2 花性状分析
17个居群的金钗石斛花形态如图3所示、花的表观指标数据如表4所示。从图3中可以直观看出不同居群金钗石斛花的形态差异明显。表4所示,花径最大的是广西金秀(GXJX)居群[(8.21±0.84) cm],最小的是西藏墨脱(XZMT)居群[(6.64±0.77) cm],两者相差1.57 cm;云南普洱(YNPE)居群的花梗最长[(5.46±0.54) cm],西藏墨脱(XZMT)居群最短[(3.26±0.18) cm],花梗长度在5 cm 以上有5个居群,4~5 cm有9个居群,4 cm以下有3个居群;唇瓣最长达到3.13 cm,为广西金秀(GXJX)居群,最短仅为2.12 cm,为西藏察隅(XZCY)居群;唇瓣最宽的也为广西金秀居群[(3.22±0.31) cm],宽度最小的为四川凉山(SCLS)居群[(1.98±0.10) cm],从表4中也可以看出,不同居群唇瓣的长宽并不呈现明显的相关性;不同居群的花瓣长度均在3.25~4.25 cm,最长的是云南保山(YNBS)居群,最短的是西藏墨脱(XZMT)居群;花瓣宽也与花瓣长没有明显的相关性,最宽的是四川西昌(SCXC)居群[(2.34±0.25) cm],最窄的是西藏察隅(XZCY)居群[(1.53±0.23) cm];萼片最长的为海南白沙(HNBS)居群[(4.55±0.25) cm],但其萼片宽最小[(0.98±0.10) cm],萼片最短的是贵州赤水(GZCS)居群[(3.58±0.35) cm],但其萼片宽最大[(1.71±0.14) cm]。
表4 金钗石斛17个居群的花形态数据
Table 4
| 编码 | 花径/cm | 花梗长/cm | 唇瓣长/cm | 唇瓣宽/cm | 花瓣长/cm | 花瓣宽/cm | 萼片长/cm | 萼片宽/cm |
|---|---|---|---|---|---|---|---|---|
| GXBS | 8.03±0.62 | 4.52±0.33 | 2.41±0.16 | 2.49±0.12 | 4.01±0.31 | 2.25±0.11 | 4.12±0.21 | 1.29±0.08 |
| GXJX | 8.21±0.84 | 5.08±0.44 | 3.13±0.24 | 3.22±0.31 | 4.16±0.22 | 2.25±0.14 | 4.36±0.33 | 1.53±0.03 |
| GXBY | 7.92±0.84 | 4.55±0.17 | 2.91±0.18 | 2.59±0.25 | 4.03±0.26 | 2.15±0.21 | 4.22±0.33 | 1.21±0.11 |
| GXRX | 7.32±0.88 | 4.64±0.18 | 2.88±0.22 | 2.46±0.20 | 3.42±0.34 | 2.18±0.30 | 3.92±0.50 | 1.18±0.12 |
| YNBS | 8.12±0.92 | 5.12±0.35 | 2.93±0.31 | 2.62±0.19 | 4.25±0.29 | 2.26±0.17 | 3.88±0.41 | 1.31±0.09 |
| YNHH | 8.20±0.78 | 4.99±0.62 | 2.68±0.28 | 2.82±0.17 | 3.97±0.54 | 2.21±0.16 | 3.64±0.32 | 1.38±0.03 |
| YNLC | 8.16±0.66 | 5.23±0.71 | 2.38±0.19 | 2.59±0.33 | 4.12±0.42 | 2.19±0.20 | 3.82±0.18 | 1.29±0.08 |
| YNPE | 8.05±0.92 | 5.46±0.54 | 2.73±0.17 | 2.56±0.21 | 3.89±0.33 | 2.27±0.22 | 4.02±0.26 | 1.48±0.09 |
| GZCS | 7.68±0.91 | 4.48±0.33 | 2.93±0.25 | 3.01±0.15 | 3.64±0.30 | 2.22±0.14 | 3.58±0.35 | 1.71±0.14 |
| GZLB | 7.75±0.75 | 4.56±0.58 | 2.95±0.31 | 2.68±0.22 | 3.59±0.18 | 2.18±0.16 | 3.67±0.34 | 1.43±0.09 |
| XZCY | 7.01±0.63 | 4.02±0.31 | 2.12±0.11 | 2.33±0.18 | 3.37±0.21 | 1.53±0.23 | 3.73±0.21 | 1.08±0.07 |
| XZMT | 6.64±0.77 | 3.26±0.18 | 2.36±0.22 | 2.28±0.12 | 3.25±0.16 | 1.78±0.13 | 3.68±0.24 | 1.25±0.12 |
| SCLS | 7.35±0.76 | 3.67±0.25 | 2.32±0.18 | 1.98±0.10 | 3.76±0.42 | 2.16±0.07 | 3.95±0.39 | 1.17±0.14 |
| SCXC | 7.73±0.89 | 3.36±0.30 | 3.08±0.15 | 2.76±0.20 | 3.59±0.15 | 2.34±0.25 | 4.12±0.12 | 1.21±0.08 |
| HNBS | 7.63±0.81 | 4.83±0.52 | 2.38±0.23 | 2.43±0.19 | 3.31±0.21 | 1.75±0.09 | 4.55±0.25 | 0.98±0.10 |
| LW | 8.05±1.03 | 5.37±0.48 | 2.98±0.20 | 3.10±0.17 | 3.96±0.31 | 1.98±0.16 | 3.62±0.41 | 1.43±0.14 |
| MD | 7.95±0.62 | 4.32±0.52 | 2.38±0.19 | 2.55±0.14 | 3.67±0.44 | 2.13±0.18 | 3.78±0.25 | 1.62±0.11 |
17个居群的金钗石斛花性状数据的变异系数如表5所示。不同性状指标数据的平均变异系数不同,其中萼片和唇瓣的性状数据平均变异系数均在8以内,最小的是唇瓣宽(7.28),萼片宽、萼片长和唇瓣长次之,花梗和花瓣性状数据的平均变异系数在8左右,平均变异系数最大的是花径(10.36)。说明金钗石斛唇瓣和萼片的性状稳定性更高,可作为优选性状指标应用于金钗石斛花性状的评价中。
表5 金钗石斛17个居群花形态数据的变异系数
Table 5
| 编码 | 花径 | 花梗长 | 唇瓣长 | 唇瓣宽 | 花瓣长 | 花瓣宽 | 萼片长 | 萼片宽 |
|---|---|---|---|---|---|---|---|---|
| GXBS | 7.72 | 7.30 | 6.64 | 4.82 | 7.73 | 4.89 | 5.10 | 6.20 |
| GXJX | 10.62 | 10.77 | 9.66 | 7.82 | 6.34 | 5.14 | 5.49 | 10.20 |
| GXBY | 10.61 | 3.74 | 6.19 | 9.65 | 6.45 | 9.77 | 7.82 | 9.09 |
| GXRX | 12.02 | 3.88 | 7.64 | 8.13 | 9.94 | 13.76 | 12.76 | 10.17 |
| YNBS | 11.33 | 6.84 | 10.58 | 7.25 | 6.82 | 7.52 | 10.57 | 6.87 |
| YNHH | 9.51 | 12.42 | 10.45 | 6.03 | 13.60 | 7.24 | 8.79 | 2.17 |
| YNLC | 8.09 | 13.58 | 7.98 | 12.74 | 10.19 | 9.13 | 4.71 | 6.20 |
| YNPE | 11.43 | 9.89 | 6.23 | 8.20 | 8.48 | 9.69 | 6.47 | 6.08 |
| GZCS | 11.85 | 7.37 | 8.53 | 4.98 | 8.24 | 6.31 | 9.78 | 8.19 |
| GZLB | 9.68 | 12.72 | 10.51 | 8.21 | 5.01 | 7.34 | 9.26 | 6.29 |
| XZCY | 8.99 | 7.71 | 5.19 | 7.73 | 6.23 | 15.03 | 5.63 | 6.48 |
| XZMT | 11.60 | 5.52 | 9.32 | 5.26 | 4.92 | 7.30 | 6.52 | 9.60 |
| SCLS | 10.34 | 6.81 | 7.76 | 5.05 | 11.17 | 3.24 | 9.87 | 11.97 |
| SCXC | 11.51 | 8.93 | 4.87 | 7.25 | 4.18 | 10.68 | 2.91 | 6.61 |
| HNBS | 10.23 | 8.66 | 7.67 | 9.63 | 5.29 | 6.22 | 7.57 | 1.96 |
| LW | 12.80 | 8.94 | 6.71 | 5.48 | 7.83 | 8.08 | 11.33 | 9.79 |
| MD | 7.80 | 12.04 | 7.98 | 5.49 | 11.99 | 8.45 | 6.61 | 6.79 |
| 均值 | 10.36 | 8.65 | 7.88 | 7.28 | 7.91 | 8.22 | 7.72 | 7.33 |
图3

图3
金钗石斛17个居群的花形态
Fig.3
Flower morphology of Dendrobium nobile in 17 populations
2.3 气孔显微结构分析
17个居群的金钗石斛植株气孔在10×40倍显微镜下观察的形态如图4所示、指标数据如表6所示。如表6所示,气孔长度最大是海南白沙(HNBS)居群(39.47 μm),最小是四川凉山(SCLS)居群(24.28 μm);气孔宽度最大是缅甸(MD)居群(34.02 μm),最小是四川西昌(SCXC)居群(16.96 μm);气孔比最大的广西金秀(GXJX)居群(1.48 μm)是最小的缅甸(MD)居群(1.14 μm)的1.30倍;缅甸(MD)居群的气孔面积最大(1 031.37 μm2),是最小的四川西昌(SCXC)居群的3.63倍;气孔密度与气孔大小基本呈反相关,气孔密度最大的是四川西昌(SCXC)居群(65.60个/mm2),气孔密度最小是海南白沙(HNBS)居群(20.80个/mm2),前者是后者的3.15倍。
图4

图4
金钗石斛17个居群的气孔显微观察(10×40倍显微镜)
Fig.4
Microscopic observation of Dendrobium nobile stomata in 17 populations (10 × 40×microscope)
表6 金钗石斛17个居群气孔的指标数据
Table 6
| 编码 | 气孔长/μm | 气孔宽/μm | 气孔比 | 气孔面积/μm2 | 气孔密度/(个/mm2) |
|---|---|---|---|---|---|
| GXBS | 30.05±1.22 | 25.06±1.18 | 1.20±0.08 | 591.15±40.15 | 35.20±2.26 |
| GXJX | 28.12±1.56 | 18.98±0.91 | 1.48±0.11 | 418.97±33.25 | 39.80±3.82 |
| GXBY | 31.34±2.24 | 24.12±1.88 | 1.30±0.09 | 593.40±25.60 | 27.60±2.48 |
| GXRX | 36.22±1.88 | 28.68±1.36 | 1.26±0.08 | 815.45±68.12 | 25.00±3.12 |
| YNBS | 32.65±1.35 | 26.18±2.21 | 1.25±0.12 | 671.00±55.85 | 36.20±3.28 |
| YNHH | 33.29±0.88 | 28.39±0.89 | 1.17±0.06 | 741.91±63.21 | 28.40±2.29 |
| YNLC | 30.31±2.21 | 24.33±0.33 | 1.25±0.08 | 578.89±50.26 | 29.80±1.95 |
| YNPE | 29.85±0.80 | 21.93±2.08 | 1.36±0.11 | 513.87±45.98 | 32.60±3.18 |
| GZCS | 25.67±1.33 | 18.66±1.09 | 1.38±0.10 | 376.02±33.28 | 51.20±3.68 |
| GZLB | 26.54±1.86 | 21.23±1.56 | 1.25±0.05 | 442.30±22.09 | 48.80±4.32 |
| XZCY | 31.56±3.12 | 25.52±0.58 | 1.24±0.04 | 632.25±48.92 | 63.60±4.59 |
| XZMT | 24.39±1.68 | 17.39±0.98 | 1.40±0.06 | 332.95±30.28 | 61.20±5.56 |
| SCLS | 24.28±1.75 | 19.33±1.21 | 1.26±0.03 | 368.43±29.85 | 63.60±5.48 |
| SCXC | 21.35±2.18 | 16.96±0.68 | 1.26±0.11 | 284.25±23.38 | 65.60±3.56 |
| HNBS | 39.47±2.58 | 32.37±0.96 | 1.22±0.13 | 1002.95±88.31 | 20.80±2.25 |
| LW | 35.09±0.95 | 30.08±3.01 | 1.17±0.05 | 828.57±65.33 | 26.40±2.30 |
| MD | 38.62±1.29 | 34.02±2.05 | 1.14±0.06 | 1031.37±81.69 | 22.60±2.20 |
17个居群金钗石斛植株气孔数据的变异系数如表7所示。不同气孔数据的平均变异系数不同,其中气孔长和宽的平均变异系数小,分别为5.74和5.58,其次为气孔比,平均变异系数为6.27,第四为气孔面积,平均变异系数为7.85,平均变异系数最大的是气孔密度(8.62)。说明气孔长和气孔宽性状稳定性高,可作为优选性状指标应用于金钗石斛气孔性状的评价中。
表7 金钗石斛17个居群气孔数据的变异系数
Table 7
| 编码 | 气孔长 | 气孔宽 | 气孔比 | 气孔面积 | 气孔密度 |
|---|---|---|---|---|---|
| GXBS | 4.06 | 4.71 | 7.43 | 7.94 | 6.42 |
| GXJX | 5.55 | 4.79 | 6.92 | 4.31 | 9.60 |
| GXBY | 7.15 | 7.79 | 6.35 | 8.35 | 8.99 |
| GXRX | 5.19 | 4.74 | 9.60 | 8.32 | 12.48 |
| YNBS | 4.13 | 8.44 | 5.13 | 8.52 | 9.06 |
| YNHH | 2.64 | 3.13 | 6.40 | 8.68 | 8.06 |
| YNLC | 7.29 | 1.36 | 8.09 | 8.95 | 6.54 |
| YNPE | 2.68 | 9.48 | 7.25 | 8.85 | 9.75 |
| GZCS | 5.18 | 5.84 | 4.00 | 4.99 | 7.19 |
| GZLB | 7.01 | 7.35 | 3.23 | 7.74 | 8.85 |
| XZCY | 9.89 | 2.27 | 4.29 | 9.09 | 7.22 |
| XZMT | 6.89 | 5.64 | 2.38 | 8.10 | 9.08 |
| SCLS | 7.21 | 6.26 | 8.73 | 8.23 | 8.62 |
| SCXC | 10.21 | 4.01 | 10.66 | 8.81 | 5.43 |
| HNBS | 6.54 | 2.97 | 4.27 | 7.88 | 10.82 |
| LW | 2.71 | 10.01 | 5.26 | 7.92 | 8.71 |
| MD | 3.34 | 6.03 | 6.67 | 6.79 | 9.73 |
| 均值 | 5.74 | 5.58 | 6.27 | 7.85 | 8.62 |
2.4 总生物碱和多糖含量分析
生物碱和多糖是金钗石斛重要的活性物质,也是种质资源品质的重要考量指标物质。表8给出了17个金钗石斛居群的总生物碱和多糖含量,总生物碱含量最高的是海南白沙(HNBS)居群[(23.71±0.74) mg/g DW],最低的是老挝(LW)居群[(4.96±0.31) mg/g DW],广西地区的金钗石斛居群普遍高于云南地区的居群,贵州地区的金钗石斛居群较高,均在17 mg/g以上,而西藏地区的居群较低,均在6.5 mg/g以下;多糖含量最高的是西藏墨脱(XZMT)居群[(0.142 5±0.008 7) g/g DW]是最低含量的广西金秀(GXJX)居群[(0.042 8±0.006 3) g/g DW]的3.33倍,表8中还可看出西藏地区的金钗石斛居群的多糖含量明显高于其他地区,其次是海南白沙(HNBS)和老挝(LW)居群,广西地区的居群间差异较大,云南地区的居群间差异较小。
表8 金钗石斛17个居群的总生物碱和多糖含量及变异系数
Table 8
| 编码 | 总生物碱 | 多糖 | ||
|---|---|---|---|---|
| 含量/(mg/g DW) | 变异系数 | 含量/(g/g DW) | 变异系数 | |
| GXBS | 11.13±0.75 | 6.74 | 0.1134±0.0055 | 4.85 |
| GXBY | 17.48±0.40 | 2.29 | 0.0928±0.0029 | 3.13 |
| GXJX | 10.74±1.03 | 9.59 | 0.0428±0.0063 | 14.72 |
| GXRX | 6.46±0.78 | 12.07 | 0.0800±0.0069 | 8.63 |
| YNBS | 8.59±0.40 | 4.66 | 0.0808±0.0061 | 7.55 |
| YNHH | 9.84±0.34 | 3.46 | 0.0814±0.0071 | 8.72 |
| YNLC | 8.45±0.42 | 4.97 | 0.0876±0.0034 | 3.88 |
| YNPE | 18.38±0.72 | 3.92 | 0.0530±0.0039 | 7.36 |
| GZCS | 19.41±1.24 | 6.39 | 0.0865±0.0069 | 7.98 |
| GZLB | 17.25±0.95 | 5.51 | 0.1030±0.0079 | 7.67 |
| XZCY | 6.30±0.24 | 3.81 | 0.1350±0.0037 | 2.74 |
| XZMT | 5.36±0.43 | 8.02 | 0.1425±0.0087 | 6.11 |
| SCLS | 5.96±0.85 | 14.26 | 0.0736±0.0054 | 7.34 |
| SCXC | 13.17±1.20 | 9.11 | 0.0864±0.0075 | 8.68 |
| HNBS | 23.71±0.74 | 3.12 | 0.1220±0.0065 | 5.33 |
| LW | 4.96±0.31 | 6.25 | 0.1135±0.0054 | 4.76 |
| MD | 12.82±0.98 | 7.64 | 0.0960±0.0062 | 6.46 |
| 均值 | — | 6.58 | — | 6.82 |
不同居群总生物碱和多糖含量的变异系数如表8所示。两种代谢产物的变异系数无显著的差异,且较小,总生物碱含量的变异系数为6.58,多糖含量的变异系数6.82。说明两种代谢产物的积累量在不同的居群中表现稳定,可作为区分金钗石斛不同居群的重要指标。
2.5 聚类分析
根据金钗石斛植株形态、花形态、气孔和代谢成分含量等数据的变异系数情况,显示植株节间长、唇瓣宽、萼片宽、气孔长、气孔宽、总生物碱含量和多糖含量在居群内性状相对稳定,以此7个性状对17个居群的金钗石斛进行系统聚类分析,结果如图5所示。当欧式距离为10时,将金钗石斛野生居群分成3组。I组包含8个居群,分别为西藏墨脱(XZMT)、四川凉山(SCLS)、广西金秀(GXJX)、四川西昌(SCXC)、广西宾阳(GXBY)、云南普洱(YNPE)、贵州赤水(GZCS)、贵州荔波(GZLB),该组内又聚成2小组,墨脱、凉山、金秀、西昌聚在一起,宾阳、普洱、赤水、荔波聚为另一小组;II组也聚集了8个居群,分别为广西容县(GXRX)、老挝(LW)、云南保山(YNBS)、西藏察隅(XZCY)、广西百色(GXBS)、云南临沧(YNLC)、云南红河(YNHH)、缅甸(MD),该组内又聚成2小组,其中缅甸居群为单独的小组,其他7个居群聚为另一小组;III组为海南白沙(HNBS)单独的一个居群,与其他居群表型形状上存在较大差异,形成了岛屿隔离效应。
图5
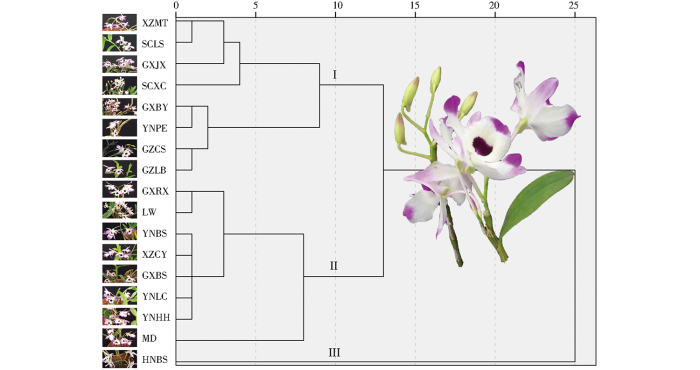
图5
金钗石斛17个居群基于性状的系统聚类分析图
Fig.5
Systematic clustering analysis based on contents of total alkaloids and polysaccharides
如表9所示,I组的8个野生金钗石斛居群表现为节短、唇瓣和萼片都较宽、气孔小,有效成分中的多糖含量低;II组的8个野生金钗石斛居群表现为节较长、气孔大,有效成分中的总生物碱含量较低;III组为海南白沙一个居群,表现为唇瓣和萼片都较窄、气孔大,有效成分中的总生物碱和多糖含量都较高。
表9 各类群金钗石斛的主要性状平均值
Table 9
| 分组 | 节间长/cm | 唇瓣宽/cm | 萼片宽/cm | 气孔长/μm | 气孔宽/μm | 总生物碱含量 /(mg/g DW) | 多糖含量 /(g/g DW) |
|---|---|---|---|---|---|---|---|
| I | 2.99 | 2.64 | 1.37 | 26.44 | 19.83 | 13.47 | 0.085 1 |
| II | 3.49 | 2.59 | 1.31 | 33.50 | 27.70 | 8.39 | 0.100 9 |
| III | 3.09 | 2.43 | 0.98 | 39.47 | 32.37 | 23.71 | 0.122 0 |
3 讨论
3.1 金钗石斛自然分布特点
金钗石斛喜欢在潮湿的环境中生长,除了少数陆生外,多为附生[12]。中国南方地处亚热带地区,亚热带季风气候是该地区的主要气候类型,其特点是气温高,夏季多雨多湿。大多数金钗石斛植物产于中国南方,因此具有高度丰富的群体。在东南亚地区,中国是金钗石斛变异性最大的国家,具有多种不同的形态特征[13]。根据本研究结果,云南省和广西壮族自治区的居群丰富度最高(各4个居群),这是因为该地区以山区为主,气候多样,包括热带山地常绿阔叶林、热带季节性潮湿林、热带季风林、热带雨林,海拔差异大,从76 m到6 800 m,该地区也是原始森林覆盖面积最大的地区,并且横断山脉阻隔了冬季的北方冷空气。此外,本研究还发现金钗石斛植物的丰富度由北向南逐渐增加,这与Pan等[14]的研究结果一致。
3.2 金钗石斛形态变异丰富
表观特性反映了植物自身的形态特点,它是植物遗传变异的直观表现,是基因调控与环境影响相互作用的结果,其优势在于可用肉眼直接将植物区分出来,遗传特征稳定,且测量方便、快捷。由于表观特性能够最为简单的反映植物的异同,因而广泛被科研学者所采用[15⇓-17]。本研究对17个居群的金钗石斛进行了表型多样性分析,发现不同居群之间的表观特性差异明显,株高较高且茎较粗的金钗石斛有云南保山、 云南临沧、贵州荔波、缅甸、老挝 5 个居群,植株矮小的有广西金秀、广西容县、西藏察隅、四川凉山4个居群,植株细长的为西藏墨脱居群,在叶片和节间长的性状上,各居群间也存在不同程度的差异。金钗石斛花型独特、颜色鲜艳,观赏价值极高[6],各居群间花的性状也存在较大差异,最大花的花径达到8.21 cm,最小花的花径仅为6.64 cm;金钗石斛花唇瓣较大的有广西金秀、贵州赤水、四川西昌和老挝居群。气孔作为植物与外界环境气体交换的通道,由两个保卫细胞构成,其形状改变直接影响气孔开闭,气孔形态特征在不同物种之间差异很大,已作为一种分类依据应用于植物分类学中[18⇓-20],气孔数量、大小、面积等在金钗石斛不同居群间差异显著,气孔面积最大达到1 031.37 μm2,最小仅为284.25 μm2;气孔密度也存在较大差异,气孔密度最大为四川西昌居群的金钗石斛,最小为海南白沙居群,两者相差3倍,通过气孔差异可以对金钗石斛进行地域区分,说明气孔性状在金钗石斛分类学中具有较高价值,气孔长宽、气孔密度及气孔面积等特征可用作金钗石斛不同居群的鉴别指标。
3.3 金钗石斛岛屿隔离与保护
中国南海作为物种迁移的屏障在塑造海南岛金钗石斛种群遗传结构方面发挥了特殊作用。持续的基因交换被认为是种群分化的主要障碍,正如 Slatkin[21]所预测,对基因流动的限制也会导致距离隔离。一个物种对岛屿栖息地的独特进化适应以及主要的水流和风向可能导致单向基因流动。这种从大陆到海岛的单向基因流可能对海南岛基因库做出了重大贡献,最终导致基因多样性增加[22-23]。金钗石斛自 1987 年起被我国列为濒危物种,保护金钗石斛野生资源的有效措施包括就地保护和异地保护。异地保护可能会减少种群间的遗传分化,由于大陆金钗石斛种群间的遗传均一性,异地保护(如收集种质资源、建立种子库、培育不同种质的金钗石斛植物等)适用于大陆金钗种群的资源保护[24]。岛屿的植物多样性指数明显高于大陆地区[25],并且由国际保护组织定义的34个生物多样性热点地区中,有20个是岛屿或由岛屿组成[26]。此外,历史上孤立的种群可能有一个独特的进化周期并可能产生新的物种,特别是由于地理隔离而在岛屿上发展出的特有类型。岛屿保存了大部分世界上受威胁生物的多样性,在岛屿上建立安全保护区已成为就地保护和遗传资源有效维护的重要保护措施[27]。因此,就地保护好我国海南岛地区金钗石斛种群是保护该物种种质资源的重要措施。
4 结论
本研究收集了17个居群215个个体的金钗石斛野生植株,对其表型性状(植株形态、花性状、气孔性状、有效成分含量)进行统计分析,并利用系统聚类分析法分析居群间的亲缘关系。结果显示,金钗石斛种质资源具有较为丰富的遗传多样性,并且根据表型性状指标将17个居群分成三组,而海南岛金钗石斛独立为一组,揭示了岛屿隔离对金钗石斛生物多样性的重要性,也为岛屿就地保护研究奠定了理论基础。
参考文献
Accurate authentication of Dendrobium officinale and its closely related species by comparative analysis of complete plastomes
Functional characterization of a novel tropinone reductase-like gene in Dendrobium nobile lindl
Effects of paclobutrazol in vitro on transplanting efficiency and root tip development of Dendrobium nobile
金钗石斛的化学成分和药理作用研究现状
Current research status of chemical constituents and pharmacological effects of Dendrobium nobile
金钗石斛的化学成分及药理作用研究进展
Research progress on chemical constituents and pharmacological effects of Dendrobium nobile
12种石斛属植物不同栽培技术及观赏价值研究
Study on different cultivation techniques and ornamental value of 12 species of Dendrobium
植物表型组学大数据及其研究进展
Big data of plant phenomics and its research progress
基于深度学习的农业植物表型研究综述
A survey on deep-learning-based plant phenotype research in agriculture
作物育种学领域新的革命: 高通量的表型组学时代
A new revolution in crop breeding: the era of high-throughput phenomics
Improving large-scale biomass and total alkaloid production of Dendrobium nobile Lindl. using a temporary immersion bioreactor system and MeJA elicitation
铁皮石斛多糖提取工艺及脱蛋白方法研究
Comparative study on extraction of crude polysaccharides from Dendrobium candidum and removing protein procedure
东南亚兰科植物的物种多样性、生活习性及其传粉系统
Species diversity, habits and pollination system of Orchidaceae in Southeast Asia
金钗石斛转录组SSR位点信息分析
SSR information in transcriptome of Dendrobium nobile
Assessing the geographical distribution of 76 Dendrobium species and impacts of climate change on their potential suitable distribution area in China
基于表型性状的甜瓜核心种质构建
Establishment of a melon (Cucumis melo) core collection based on phenotypic characters
表型水平上检验水稻核心种质的参数比较
Comparison of parameters for testing the rice core collection in phenotype
粒用高粱种质资源主要农艺性状的相关性及主成分分析
Correlation and principal component analysis of main agronomic characters of grain Sorghum germplasm resources
花楸属3种植物的基因组大小与叶气孔特征分析
Variation in genome size and stomatal traits among three Sorbus species
百部科植物叶表皮特征及其分类学意义
Characters of the leaf epidermis of Stemonaceae and their taxonomical significance
15种独蒜兰属植物叶表皮微形态特征及分类学意义研究
Leaf epidermal micro-morphology and taxonomic significance of 15 species of Pleione
Isolation by distance in equilibrium and non-equilibrium populations

It is shown that for allele frequency data a useful measure of the extent of gene flow between a pair of populations is M∘=(1/FST-1)/4, which is the estimated level of gene flow in an island model at equilibrium. For DNA sequence data, the same formula can be used if F is replaced by N. In a population with restricted dispersal, analytic theory shows that there is a simple relationship between M̂ and geographic distance in both equilibrium and non-equilibrium populations and that this relationship is approximately independent of mutation rate when the mutation rate is small. Simulation results show that with reasonable sample sizes, isolation by distance can indeed be detected and that, at least in some cases, non-equilibrium patterns can be distinguished. This approach to analyzing isolation by distance is used for two allozyme data sets, one from gulls and one from pocket gophers.© 1993 The Society for the Study of Evolution.
van oosterhout C. Population genetic analysis of microsatellite variation of guppies (Poecilia reticulata) in Trinidad and Tobago: evidence for a dynamic source-sink metapopulation structure, founder events and population bottlenecks
Riverine fish populations are traditionally considered to be highly structured and subject to strong genetic drift. Here, we use microsatellites to analyse the population structure of the guppy (Poecilia reticulata), focussing on the headwater floodplain area of the Caroni drainage in Trinidad. We also analyse the population genetics of guppies in the Northern Drainage in Trinidad, a habitat characterized by rivers flowing directly into the sea, and a small isolated population in Tobago. Upland Caroni populations are highly differentiated and display low levels of genetic diversity. However, we found no evidence to suggest that these upland populations experienced recent population crashes and the populations appear to approach mutation-drift equilibrium. Dominant downstream migration over both short- and long-time frames has a strong impact on the population genetics of lowland Caroni populations. This drainage system could be considered a source-sink metapopulation, with the tributary furthest downstream representing a 'super sink', receiving immigrants from rivers upstream in the drainage. Moreover, the effective population size in the lowlands is surprisingly low in comparison with the apparently large census population sizes.
Multiscale analysis of Hymenocallis coronaria (Amaryllidaceae) genetic diversity, genetic structure, and gene movement under the influence of unidirectional stream flow
Understanding gene movement patterns in unidirectional flow environments and their effect on patterns of genetic diversity and genetic structure is necessary to manage these systems. Hypotheses and models to explain genetic patterns in streams are rare, and the results of macrophyte studies are inconsistent. This study addresses Ritland's (Canadian Journal of Botany 67: 2017-2024) unidirectional diversity hypothesis, the one-dimensional stepping stone model, and the metapopulation model within and among populations. Hymenocallis coronaria, an aquatic macrophyte of rocky river shoals of the SE USA, was sampled in four river basins. Within populations and among populations <16.2 km apart had significant isolation by distance. However, the rate of gene flow decay was not consistent with a one-dimensional stepping stone model, nor was evidence strong or consistent for Ritland's hypothesis. Some evidence indicates that localized metapopulation processes may be affecting genetic diversity and structure; however, gene flow patterns inconsistent with the assumptions of the linear and unidirectional models are also a possible influence. We discuss three variants on the one-dimensional stepping stone model. Future research in linear environments should examine the expectations of these models. This study is also one of the first efforts to calculate population genetic parameters using a new program, TETRASAT.
Genetic diversity assessment and ex situ conservation strategy of the endangered Dendrobium officinale (Orchidaceae) using new trinucleotide microsatellite markers
Different evolutionary processes in shaping the genetic composition of Dendrobium nobile in southwest China
Conservation of oceanic island floras: present and future global challenges

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}